(Like the title? I do a love a touch of alliteration…) It’s been a while since I wrote, but our lab group have a couple of recent publications out based on scanning of stem-tetrapod (tetrapodomorph fish) fossils that I’d love to fill you in on.
Firstly, PhD candidate Corinne Mensforth led a study on an enigmatic fish from Antarctica called Koharalepis. This species was first described by Gavin Young and colleagues in the 90s, and in 2018, John Long, Brian Choo and I published a paper based on synchrotron scans of the holotype and only known specimen. However, this was a difficult specimen to image and it had kept many of it’s secrets from us … until now.

More recently, we have used neutron tomography to scan this fish, a technique which has finally revealed to us much more of the internal features preserved within this fossil that we had been able to see before. This led to Corinne’s paper, “New data on the sarcopterygian Koharalepis jarviki (Tetrapodomorpha; Canowindridae) from the Late Devonian of Antarctica, revealed via synchrotron and neutron tomography” published in the journal Frontiers in Ecology and Evolution in April this year.
It was great to see Koharalepis get a moment in the spotlight, with the media release getting picked up all over the world, including a mention in IFLScience! This was certainly helped by the stunning life reconstruction that our talented Honours student, Tom Turner, created. Just beautiful!!

NEXT, and keeping on with the theme of high powered scanning of stem-tetrapod fossils, today my long-awaited publication “Ethmosphenoid endocasts elucidate evolutionary braindivergences and interrelationships of stem tetrapods (Sarcopterygii, Tetrapodomorpha)” was published in Papers in Palaeontology, with a co-author list that truly spans the globe. Big thanks to all my co-authors, Richard Cloutier, Oleg Lebedev, Tom Challands, Jing Lu, Joseph Bevitt, Laurent Houle, Shaun Collin, & John Long. This one took a team effort and a long time…

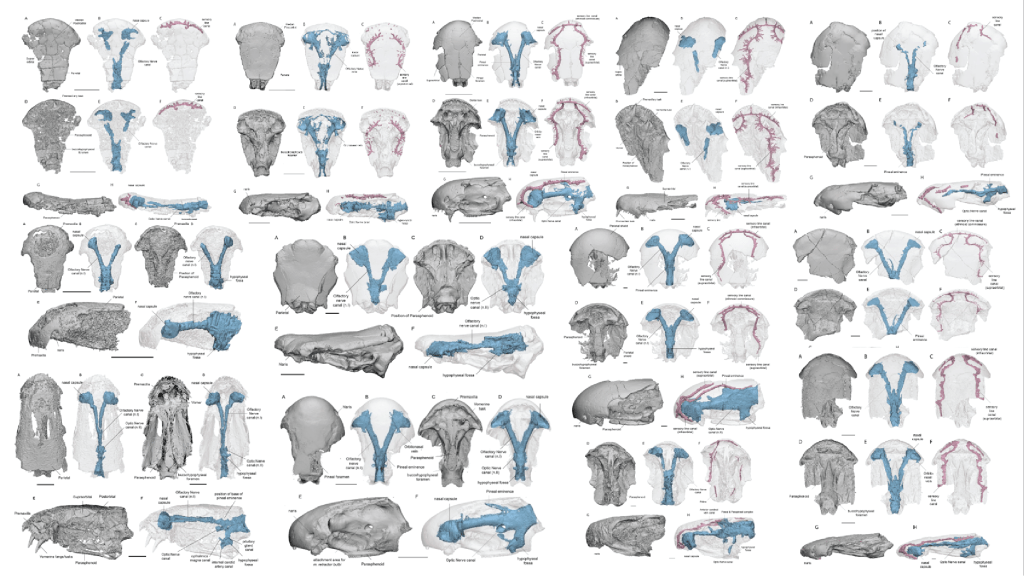
In this work we scaned 13 taxa of stem tetrapods across three families (‘Tristichopteridae’, ‘Osteolepididae’, Megalichthyidae) to examine skull and endocast morphology using Principal Components Analysis. Of note there are some particularly mysterious and otherwise poorly-known species included in this analysis, including some more from Antarctica (Mahalalepis, Platyethmoidia, and a new still unamed fossil), Australia (Owensia), and Russia (Megapomus), alongside other better known species such as Gogonasus and Eusthenopteron.
Our study shows that the internal cranial endocast can be useful to help identify phylogentic affinity (relationships to other species) in forms where that is otherwise difficult to discern. We also deduce that stem tetrapods were likely quite conservative in their palaeoneurology (brain shape), in contrast to what we see in lungfishes, for example.
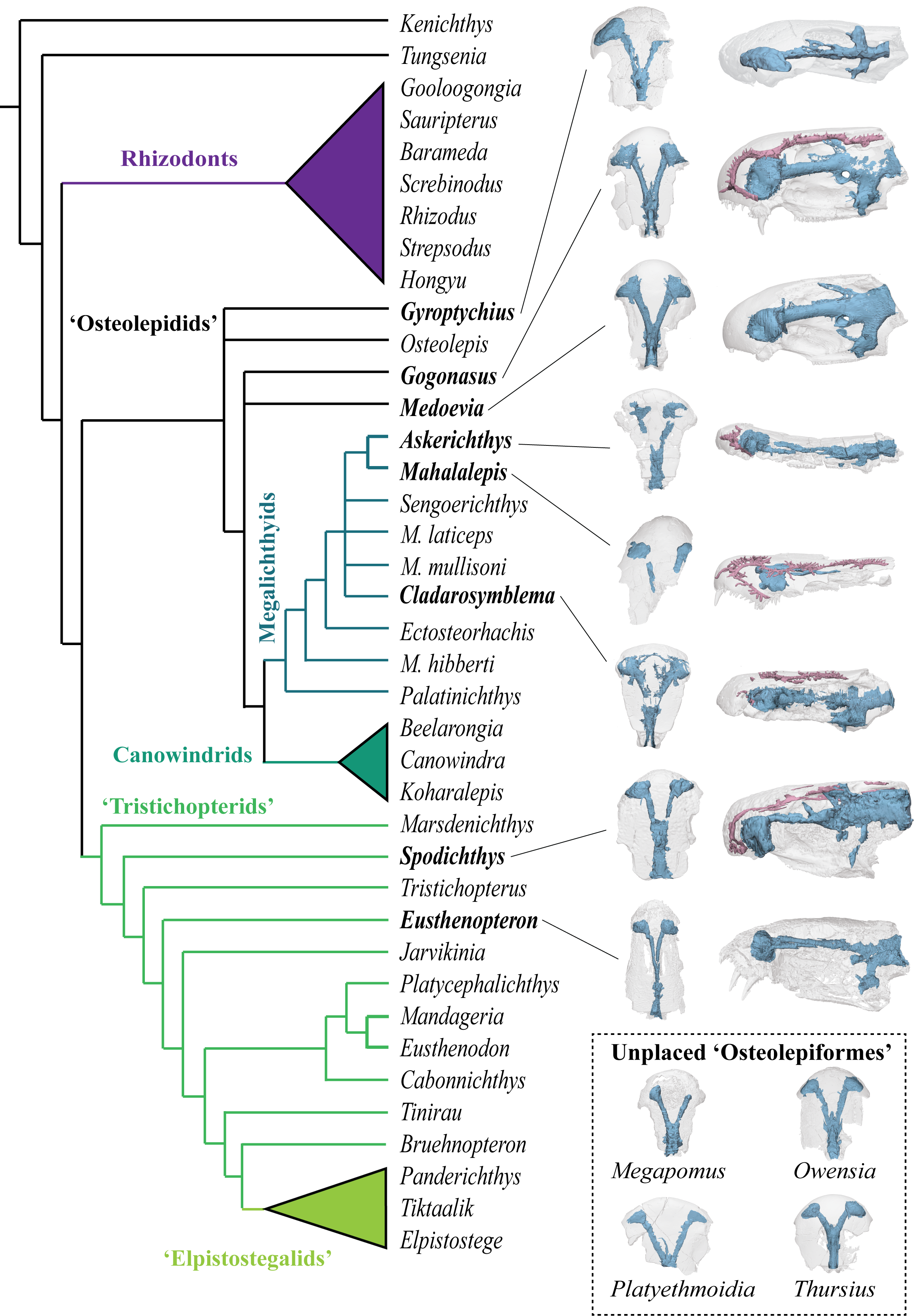
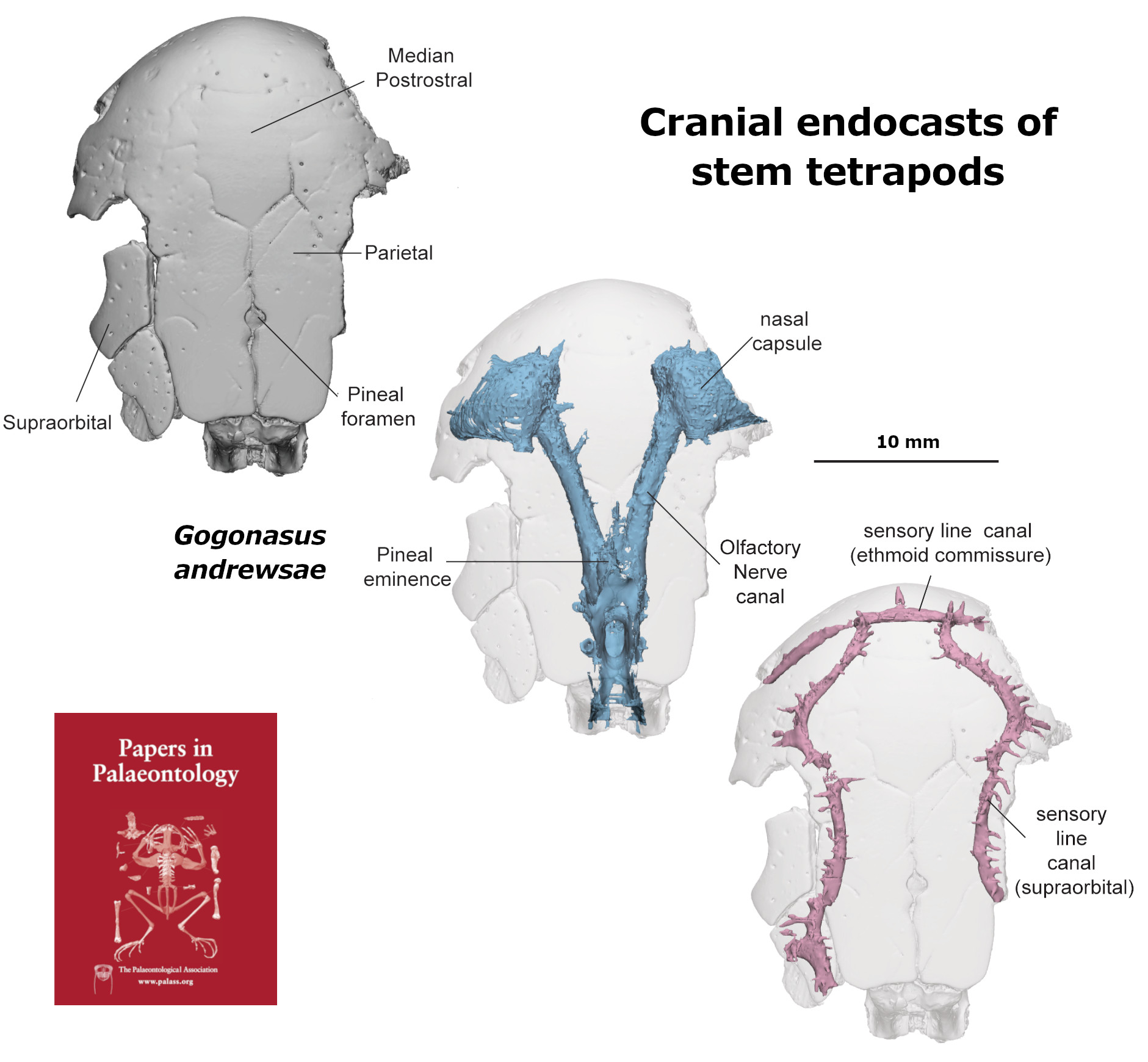
The scanning data and resultant 3D digital models for both studies are online and can be viewed via their respective projects on MorphoSource: Koharalepis, and Ethmosphenoid endocasts of stem tetrapods.